
Bioenergetics: Energy Transformation in Living Systems
An IMAT Review on How Living Organisms Manage Their Energy Resources
Introduction to Bioenergetics
Bioenergetics is the branch of biology that studies how living organisms manage their energy resources. This involves examining the intricate processes by which energy is captured, converted from one form to another, stored, and utilized to sustain life. At its core, bioenergetics focuses on the transformation of energy. All biological activities, from the microscopic level of molecular synthesis and active transport across cell membranes to the macroscopic level of muscle contraction and nerve impulse transmission, are fundamentally dependent on a continuous supply and regulated expenditure of energy. Understanding these energy conversion processes is not merely an academic exercise; it is fundamental to comprehending the very essence of life, forming a critical foundation for fields such as medicine and biotechnology.
The various topics within bioenergetics, such as the role of adenosine triphosphate (ATP), the principles of metabolism, the function of enzymes, and the complex pathways of cellular respiration and photosynthesis, are not isolated concepts. Instead, they represent deeply interconnected components of a cell's comprehensive energy strategy. ATP serves as the universal energy currency, readily available to power cellular work. Metabolism, encompassing both catabolic (breakdown) and anabolic (synthesis) reactions, dictates the flow of this energy – catabolism generates ATP, while anabolism consumes it. Enzymes are the biological catalysts that orchestrate these metabolic reactions, ensuring they occur at rates compatible with life. Cellular respiration is a prime example of a catabolic pathway, efficiently extracting energy from fuel molecules like glucose to produce ATP. Conversely, photosynthesis, in organisms capable of it, is a major anabolic pathway that captures external light energy to synthesize glucose, which subsequently fuels respiration. Recognizing these interconnections is crucial for a holistic understanding of how cells and organisms thrive by dynamically balancing energy supply and demand.
🔬 I. ATP: The Universal Energy Currency
A. Structure of Adenosine Triphosphate (ATP)
ATP is classified as a nucleoside triphosphate. This classification is significant as it links ATP structurally to the building blocks of nucleic acids (RNA and DNA) and to various signaling molecules within the cell. The molecule itself is composed of three distinct parts:
- Adenine: A nitrogenous base, specifically a purine, which provides part of the molecule's identity.
- Ribose: A five-carbon sugar (a pentose) that is attached to the adenine base. Together, adenine and ribose form the nucleoside called adenosine.
- Three Phosphate Groups: These are linked in series to the ribose sugar. The bonds connecting these phosphate groups, particularly the bonds between the second and third phosphate groups (known as phosphoanhydride bonds), are crucial for ATP's energy-carrying function.

📸 Source/Description: The chemical structure of ATP, clearly labeling the adenine base, the ribose sugar, and the three phosphate groups. Arrows would point to the high-energy phosphoanhydride bonds between the phosphate groups.
The term "high-energy bonds" associated with ATP can sometimes be misconstrued. It is not that these specific bonds are inherently stronger than other covalent bonds. Rather, the energy release upon hydrolysis of these bonds is substantial because the products of this reaction (adenosine diphosphate, ADP, and inorganic phosphate, Pᵢ) are significantly more stable and at a lower energy state than the ATP molecule itself. The three phosphate groups in ATP are negatively charged and are held in close proximity, leading to considerable electrostatic repulsion, much like a compressed spring. When ATP is hydrolyzed to ADP and Pᵢ, one of these phosphate groups is removed, relieving some of this repulsion. This transition to a more stable, lower-energy state results in the release of a significant amount of free energy (approximately -7.3 kcal/mol or -30.5 kJ/mol under standard conditions), which the cell can then harness for work.
B. ATP's Role in Cellular Processes
ATP's designation as the "energy currency" stems from its ability to provide readily releasable energy to power a vast array of cellular functions. Just as currency facilitates economic transactions, ATP facilitates energy transactions within the cell. Its hydrolysis to ADP and Pᵢ is coupled to endergonic (energy-requiring) reactions, making them thermodynamically favorable.
Key cellular processes fueled by ATP include:
- Mechanical Work: Such as the contraction of muscle fibers, movement of cilia and flagella, and the movement of chromosomes during cell division.
- Transport Work: The active transport of ions and molecules across cell membranes against their concentration gradients. A classic example is the sodium-potassium pump, essential for maintaining cellular membrane potential and nerve function.
- Chemical Work: The synthesis of complex molecules (anabolism), such as the formation of polymers like proteins from amino acids and nucleic acids from nucleotides. This also includes substrate phosphorylation, where ATP donates a phosphate group to another molecule, often activating it for a subsequent reaction.
- Signaling: ATP itself can act as an extracellular signaling molecule by binding to purinergic receptors on cell surfaces. Furthermore, its derivative, cyclic AMP (cAMP), formed from ATP by the enzyme adenylate cyclase, is a crucial intracellular second messenger involved in many signal transduction pathways.
- Genetic Material Synthesis: ATP is one of the four nucleotide triphosphates required as a monomer for the synthesis of RNA. The energy for DNA synthesis is also derived from the hydrolysis of deoxyribonucleotide triphosphates, which are structurally analogous to ATP.
The versatility of ATP extends beyond its role as a direct energy donor. Its involvement in signaling pathways and its function as a direct precursor for RNA synthesis highlight how a single molecular structure can be adapted for multiple critical cellular functions. This molecular multitasking is a common theme in biochemistry, reflecting the efficiency of biological design. Such broader understanding of molecular roles is important for comprehensive biological knowledge.
C. The ATP-ADP Cycle: Recharging the Cellular Battery
Cells maintain a dynamic balance of ATP through the continuous ATP-ADP cycle, which can be likened to the charging and discharging of a rechargeable battery.
Hydrolysis (Discharge): When the cell requires energy for work, ATP undergoes hydrolysis. An enzyme (typically an ATPase) catalyzes the reaction. This exergonic reaction releases energy that the cell can use. ADP (adenosine diphosphate) and an inorganic phosphate group (Pᵢ) are the products.
Phosphorylation/Synthesis (Recharge): To replenish ATP, ADP is re-phosphorylated to ATP. This endergonic reaction requires an input of energy, which is typically derived from catabolic processes, primarily cellular respiration (the breakdown of glucose and other fuel molecules) or, in photosynthetic organisms, from light energy captured during photosynthesis. Enzymes like ATP synthase are crucial for this process.
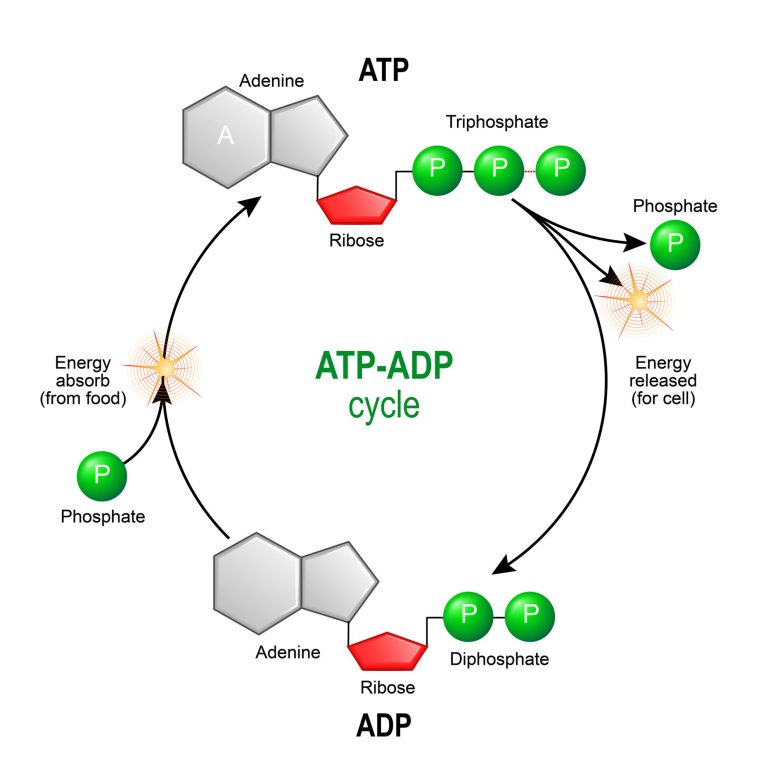
📸 Source/Description: This figure depicts the cyclical process where ATP is converted into ADP via hydrolysis (one phosphate group being removed), and then ADP is converted back into ATP via phosphorylation (one phosphate group being added), fueled by cellular processes.
Cells do not maintain vast stockpiles of ATP. Instead, ATP is constantly being used and regenerated. The intracellular concentration of ATP is relatively low, and a typical ATP molecule is consumed and regenerated very rapidly, often within a minute. This high turnover rate underscores the cell's continuous high demand for energy and the efficiency of the ATP-ADP cycle. If ATP were a very stable molecule stored in large quantities, its energy would not be "readily releasable" for immediate needs. Its moderate instability and rapid turnover make it an exceptionally efficient transient carrier of energy, perfectly suited to the dynamic energy requirements of living cells. This rapid cycling ensures a constant supply of energy, allowing cells to respond quickly to changing metabolic demands.
🔄 II. Metabolism: The Sum of Cellular Reactions
Metabolism refers to the entirety of chemical reactions that occur within a living organism to maintain life. These reactions are organized into metabolic pathways, which are sequences of enzyme-catalyzed steps. Metabolism can be broadly divided into two opposing but interconnected processes: anabolism and catabolism.
A. Anabolism: Building Up Molecules (Energy Input)
Anabolic pathways, also known as biosynthetic pathways, involve the construction of complex molecules from simpler precursors. These processes are fundamental for growth, repair, and the storage of energy. Anabolic reactions are endergonic, meaning they require an input of energy to proceed. This energy is typically supplied by the hydrolysis of ATP.
Examples of anabolic processes include:
- Photosynthesis: In plants and some other organisms, the synthesis of glucose from carbon dioxide and water using light energy.
- Protein synthesis: The formation of proteins from amino acids.
- DNA replication: The synthesis of DNA from deoxyribonucleotides.
- Glycogen synthesis: The formation of glycogen (a storage form of glucose) from glucose molecules.
B. Catabolism: Breaking Down Molecules (Energy Release)
Catabolic pathways, also known as degradative pathways, involve the breakdown of complex molecules into simpler ones. These processes release energy, some of which is captured in the form of ATP, while the rest is lost as heat. Catabolic reactions are exergonic, meaning they release energy.
Examples of catabolic processes include:
- Cellular respiration: The breakdown of glucose and other fuel molecules (like fatty acids) in the presence of oxygen to produce carbon dioxide, water, and a significant amount of ATP. Glycolysis and the Citric Acid Cycle (Krebs Cycle) are key components of this catabolic process.
- Digestion: The breakdown of large food molecules (carbohydrates, proteins, fats) into smaller absorbable units in the digestive tract.
- Breakdown of stored reserves: Such as the breakdown of glycogen to glucose or fats to fatty acids and glycerol when energy is needed.
C. Interplay between Anabolism and Catabolism
Anabolism and catabolism are not independent processes; they are intricately linked and carefully regulated to meet the cell's needs. The energy released during catabolic reactions is harnessed to drive anabolic reactions, with ATP serving as the primary energy-carrying molecule that couples these two arms of metabolism. Catabolic pathways provide the ATP and the simple precursor molecules (building blocks) that anabolic pathways require.
Enzymes are crucial for controlling the rate and direction of all metabolic pathways, both anabolic and catabolic. Many metabolic pathways are also reversible, allowing cells to adapt to changing conditions and substrate availability. This dynamic interplay ensures that the cell can efficiently manage its resources, building necessary components when energy and materials are available, and breaking down stores when energy is required. The concept of energy coupling via ATP is fundamental: thermodynamically unfavorable anabolic reactions are made possible by linking them to the highly favorable hydrolysis of ATP, which itself is generated by favorable catabolic reactions. This elegant system underpins all cellular activity.
| Comparison of Anabolism and Catabolism | ||
|---|---|---|
| Feature | Anabolism | Catabolism |
| Definition | Building complex molecules from simpler ones | Breaking down complex molecules into simpler ones |
| Energy | Requires energy input (endergonic) | Releases energy (exergonic) |
| Type of Reaction | Synthesis (biosynthesis) | Degradation |
| Role of ATP | Consumes ATP | Generates ATP |
| Examples | Protein synthesis, Photosynthesis, DNA replication | Cellular respiration, Digestion, Glycogenolysis |
| Effect on Molecular Size | Increases molecular size and complexity | Decreases molecular size and complexity |
⚙️ III. Enzymes: Facilitators of Life's Reactions
A. Enzyme Structure and the Active Site
Enzymes are vital biological molecules, predominantly proteins, that act as catalysts to accelerate the rate of biochemical reactions within cells. They achieve this without being consumed or permanently altered in the process. Virtually every metabolic reaction is facilitated by a specific enzyme, making them indispensable for life.
The primary structure of an enzyme, which is the linear sequence of its amino acids, dictates its unique three-dimensional conformation. This specific 3D structure is crucial for its function, particularly a region known as the active site. The active site is a small pocket or crevice on the enzyme's surface where the reactant molecules, called substrates, bind and where the chemical reaction takes place. The shape, size, and chemical properties (e.g., charge distribution, hydrophobicity) of the active site are precisely tailored to bind specific substrates, conferring high specificity to enzyme action.
Some enzymes, termed apoenzymes, are inactive in their protein-only form. They require the binding of a non-protein component called a cofactor to become active. Cofactors can be inorganic metal ions (e.g., Zn2+, Mg2+) or complex organic molecules known as coenzymes (often derived from vitamins, e.g., NAD+, FAD). When an apoenzyme is bound to its necessary cofactor(s), the complete, active enzyme is called a holoenzyme.
B. How Enzymes Work: Substrate Specificity
Enzymes function by lowering the activation energy () of a reaction. Activation energy is the minimum energy required for a chemical reaction to occur. By reducing this energy barrier, enzymes significantly increase the rate at which the reaction proceeds, often by factors of millions or more. It is important to note that enzymes do not alter the overall free energy change () or the equilibrium position of a reaction; they only affect the rate at which equilibrium is reached.
_DRAFT_Two main models describe the interaction between an enzyme and its substrate:
- Lock-and-Key Model: Proposed by Emil Fischer, this earlier model suggests that the active site of an enzyme has a rigid shape that is precisely complementary to the shape of its substrate, much like a specific key fits into a particular lock. This model effectively explains the high specificity of many enzymes for their substrates. When the substrate enters the active site, it fits perfectly, forming an enzyme-substrate complex.
- Induced-Fit Model: This more contemporary model, proposed by Daniel Koshland, refines the lock-and-key concept. It suggests that the active site is not initially a perfect fit for the substrate. Instead, the binding of the substrate induces a conformational change in the enzyme's active site, leading to a more precise and tighter fit. This dynamic interaction can also strain the bonds within the substrate molecule, contorting it into its transition state and thereby facilitating the reaction. The induced-fit model offers advantages over the lock-and-key model as it can explain how some enzymes exhibit broader specificity (e.g., lipase can bind to a variety of lipid substrates) and provides a better understanding of the catalytic mechanism itself, where the conformational change actively participates in catalysis.

📸 Source/Description: The induced-fit model shows the enzyme's active site changing shape to accommodate the substrate, a refinement of the older lock-and-key theory.
The development from the lock-and-key model to the induced-fit model illustrates how scientific understanding evolves with new evidence and more sophisticated experimental techniques. While the lock-and-key model provided a foundational understanding of enzyme specificity, the induced-fit model offers a more dynamic and accurate picture of enzyme-substrate interactions, better explaining the flexibility and catalytic power of these remarkable protein machines.
C. Factors Affecting Enzyme Reaction Rates
The rate of an enzyme-catalyzed reaction can be influenced by several environmental factors:
Temperature: Generally, increasing the temperature increases the rate of an enzyme-catalyzed reaction due to increased kinetic energy. Each enzyme has an optimal temperature (around 37°C for most human enzymes). Temperatures significantly above the optimum cause the enzyme to denature, a loss of structure leading to a rapid loss of activity.
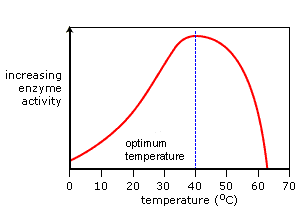
📸 Source/Description: This graph shows that enzyme activity increases with temperature to an optimal point, after which it rapidly declines due to denaturation.
pH: Each enzyme has an optimal pH range. The pH affects the ionization state of amino acid residues in the active site. Extreme pH values cause denaturation. For example, pepsin (stomach) functions optimally at pH ~2, whereas trypsin (small intestine) has an optimum pH of about 8.

📸 Source/Description: This bell-shaped curve shows that enzymes have an optimal pH. Activity decreases as the pH deviates from this optimum.
Substrate and Enzyme Concentration: Increasing substrate concentration increases the reaction rate until the enzyme is saturated (all active sites are occupied), at which point the rate reaches its maximum (Vmax) and plateaus. If substrate is in excess, increasing enzyme concentration increases the rate proportionally.
The high degree of specificity of enzymes to particular environmental conditions, such as temperature and pH, underscores their delicate nature. This sensitivity highlights the critical importance of homeostasis – the maintenance of stable internal conditions – for life.
⚡ IV. Cellular Respiration: Harvesting Energy from Glucose
A. Overview: The Big Picture
Cellular respiration is the primary catabolic process by which living cells extract chemical energy stored in organic molecules, predominantly glucose, and convert it into a usable form, ATP. In the presence of oxygen (aerobic respiration), the overall chemical equation can be summarized as:
Cellular respiration occurs in several distinct stages: Glycolysis (cytoplasm), The Krebs Cycle (mitochondrial matrix), and the Electron Transport Chain (inner mitochondrial membrane). The structure of the mitochondrion, with its double membrane and folded cristae, is crucial for the efficiency of this process.
.svg)
📸 Source/Description: Glycolysis occurs in the cytoplasm, breaking glucose into pyruvate. Pyruvate enters the mitochondrion for the Krebs Cycle and the Electron Transport Chain, which produces the majority of ATP.
B. Stage 1: Glycolysis (Cytoplasm)
Glycolysis ("sugar splitting") is an anaerobic sequence of ten reactions in the cytoplasm. One 6-carbon glucose molecule is converted into two 3-carbon pyruvate molecules. The net energy yield per glucose is 2 ATP and 2 NADH.
| Inputs and Outputs of Glycolysis (per molecule of glucose) | |
|---|---|
| Inputs | Outputs |
| 1 Glucose | 2 Pyruvate |
| 2 ATP | 4 ATP (Net 2 ATP) |
| 2 NAD+ | 2 NADH + 2 H+ |
| 4 ADP + 4 Pᵢ | 2 H₂O |
C. Stage 2: The Krebs Cycle (Citric Acid Cycle) (Mitochondrial Matrix)
If oxygen is present, pyruvate enters the mitochondrial matrix and is converted to acetyl-CoA. The Krebs cycle then completely oxidizes the acetyl group. For each glucose molecule (2 turns of the cycle), it produces 4 CO₂, 2 ATP (or GTP), 6 NADH, and 2 FADH₂.
| Inputs and Outputs of the Krebs Cycle (per molecule of glucose) | |
|---|---|
| Inputs | Outputs |
| 2 Acetyl-CoA | 4 CO₂ |
| 2 ADP + 2 Pᵢ | 2 ATP (or GTP) |
| 6 NAD+ | 6 NADH + 6 H+ |
| 2 FAD | 2 FADH₂ |
D. Stage 3: Electron Transport Chain (ETC) and Oxidative Phosphorylation
Electron Transport Chain (ETC): High-energy electrons from NADH and FADH₂ are passed down a series of protein complexes on the inner mitochondrial membrane. The energy released is used to pump protons (H+) into the intermembrane space, creating a proton-motive force. Oxygen is the final electron acceptor, forming water.
Oxidative Phosphorylation (Chemiosmosis): Protons flow back into the matrix through ATP synthase, driving the synthesis of the majority of ATP (~28-30 molecules per glucose). This process couples the oxidation of electron carriers to the phosphorylation of ADP.
E. Aerobic vs. Anaerobic Respiration
The presence or absence of oxygen dictates the metabolic pathway after glycolysis.
| Comparison of Aerobic and Anaerobic Respiration | ||
|---|---|---|
| Feature | Aerobic Respiration | Anaerobic Respiration (Fermentation) |
| Oxygen Requirement | Yes (essential) | No |
| Main Stages | Glycolysis, Krebs Cycle, ETC | Glycolysis, Fermentation |
| Final Electron Acceptor | Oxygen (O₂) | Pyruvate |
| ATP Yield (per glucose) | High (~30-32 ATP) | Low (2 ATP) |
| End Products (humans) | CO₂, H₂O, ATP | Lactic acid, ATP |
Glycolysis is a remarkably ancient metabolic pathway. Anaerobic pathways like fermentation are crucial for regenerating NAD+ from NADH, allowing glycolysis to continue in the absence of oxygen. This provides a rapid but inefficient source of ATP, which is advantageous during sudden, high energy demands like sprinting.
☀️ V. Photosynthesis: Converting Light Energy to Chemical Energy
A. Overview: The Process and Its Importance
Photosynthesis is a vital anabolic process by which plants, algae, and some bacteria convert light energy into chemical energy stored in glucose. In eukaryotes, it takes place in chloroplasts. The overall equation is:
Chloroplasts contain thylakoids (where light-dependent reactions occur) and stroma (where the Calvin cycle occurs).

📸 Source/Description: This diagram shows the two stages of photosynthesis. The light-dependent reactions in the thylakoids produce ATP and NADPH, which are used in the Calvin cycle in the stroma to fix CO₂ into sugar.
B. Light-Dependent Reactions (Thylakoid Membranes)
These reactions capture light energy to produce ATP and NADPH. Key steps include light absorption by photosystems, splitting of water (photolysis), an electron transport chain, and the formation of ATP and NADPH.
| Inputs and Outputs of Light-Dependent Reactions | |
|---|---|
| Inputs | Outputs |
| Light Energy | ATP |
| Water (H₂O) | NADPH |
| ADP + Pᵢ | Oxygen (O₂) |
| NADP+ | |
C. Light-Independent Reactions (Calvin Cycle) (Stroma)
The Calvin cycle uses ATP and NADPH to convert atmospheric CO₂ into sugar. It has three phases: Carbon Fixation (catalyzed by RuBisCO), Reduction, and Regeneration of the starting molecule (RuBP).
| Inputs and Outputs of the Calvin Cycle (per 1/2 glucose) | |
|---|---|
| Inputs | Outputs |
| 3 CO₂ | 1 G3P (Glyceraldehyde-3-phosphate) |
| 9 ATP | 9 ADP + 8 Pᵢ |
| 6 NADPH | 6 NADP+ |
D. Factors Affecting the Rate of Photosynthesis
The rate of photosynthesis is influenced by limiting factors. The rate is governed by the factor in the least optimal amount.
- Light Intensity: Rate increases with light until a saturation point is reached.
- Carbon Dioxide Concentration: Rate increases with CO₂ until a saturation point is reached.
- Temperature: Rate increases to an optimal temperature, then drops sharply as enzymes denature.
📸 Source/Description: These graphs illustrate how light intensity, temperature, and CO₂ concentration can each act as a limiting factor on the rate of photosynthesis.
🌍 Conclusion: Interconnectedness of Bioenergetic Pathways
The processes of cellular respiration and photosynthesis are cornerstones of bioenergetics, representing the primary mechanisms by which energy is transformed and utilized by living organisms. Cellular respiration meticulously extracts energy from organic molecules, primarily glucose, converting it into ATP, the cell's universal energy currency. Photosynthesis, conversely, captures light energy and stores it in the chemical bonds of glucose, forming the base of most food chains and replenishing the atmospheric oxygen vital for aerobic life.
ATP stands as the central intermediary in cellular energy transactions, effectively linking catabolic pathways (like respiration, which release energy and produce ATP) with anabolic pathways (like the synthesis of complex molecules and aspects of photosynthesis, which require energy and consume ATP). This coupling ensures that energy released from fuel breakdown can be efficiently directed towards life-sustaining work.
None of these intricate metabolic transformations would be possible at rates compatible with life without the action of enzymes. These biological catalysts accelerate reactions by lowering activation energies and are themselves subject to regulation, allowing cells to fine-tune their metabolic activities in response to changing needs and environmental conditions. The sensitivity of enzymes to factors like temperature and pH underscores the importance of homeostasis for maintaining optimal cellular function.
Furthermore, there is a profound cyclical relationship in the flow of matter and energy within ecosystems, largely driven by these bioenergetic pathways. Photosynthesis consumes carbon dioxide and water and releases oxygen and glucose. Cellular respiration consumes oxygen and glucose and releases carbon dioxide and water. This grand cycle sustains life on Earth, highlighting the interdependence of organisms and their environment.
Ultimately, the efficient capture, conversion, storage, and utilization of energy, as detailed through the study of bioenergetics, are fundamental prerequisites for the complexity, order, and responsiveness that characterize living systems. A thorough understanding of bioenergetics is therefore not just about memorizing pathways and molecules; it is about appreciating the dynamic and exquisitely regulated flow of energy that makes life possible.